Introduction: The Central Role of Carbohydrates
Carbohydrates are a class of molecules that play a foundational role in biology and are, therefore, a critical topic for the Medical College Admission Test (MCAT). These :hydrated carbons” serve as the principal energy source for cellular metabolism, a readily accessible storage for of energy (glycogen in animals, starch in plants), and a key structural component in organisms (cellulose in plants). A deep understanding of their structure and chemistry is essential to grasp why we can digest potatoes and cotton candy for energy but not wood or a cotton T-shirt, despite all being digest composed of glucose. The key lies in the specific linkages between sugar units, a distinction that determines their biological function and digestibility. This guide provides a comprehensive, integrated review of carbohydrate structure, chemistry, and the metabolic pathways that govern their use, providing the foundational knowledge necessary for success on the MCAT.
1. The Building Blocks – Monosaccharides
A mastery of monosaccharide structure and classification is the non-negotiable foundation for understanding all carbohydrate metabolism. These simple sugars are the fundamental building blocks of more complex carbohydrates and the primary cyrrency of the metabolic pathways discussed later in this guide. The following sections will walk you through the fundamental chemistry, stereochemistry, and variations of these single sugar units.
1.1 Fundamental Structure and Classification
A monosaccharide, or simple sugar, is defined by its chemical composition and core structural features. They adhere to the general formula (CH₂O)ₙ, signifying that they are composed of carbons hydrated with water. Structurally, they consist of an uninterrupted carbon chain where one carbon is a carbonyl group (either an aldehyde or a ketone) and all other carbons bear an alcohol (hydroxyl) group.
Monosaccharides are classified using two primary methods, which are often combined to create a more descriptive name.
| Classification by number of Carbons | Classification by Functional Group |
|---|---|
| Triose (3 carbons) | Aldose: contains an aldehyde functional group. The carbonyl carbon (anomeric carbon) is designated carbon 1 (C1) |
| Tetrose (4carbons) | Ketose: Contains a ketone functional group. In biologically relevant ketoses, the anomeric carbon is typically found at carbon 2 (C2) |
| Pentose (5 carbons), e.g., Ribose | |
| Hexose (6 carbons), e.g., Glucose, Fructose, Galactose |
Using the combined classification system, we can be more specific. For example, D-glucose, which has six carbons and an aldehyde group, is an aldohexose. In contrast, ribulose, with five carbons and a ketone group, is a ketopentose.
1.2 Stereochemistry of Monosaccharides
Monosaccharides with the same molecular formula can exist in different structural forms. Constitutional isomers have the same formula but different connectivity; for instance, glucose (an aldose) and fructose (a ketose) are constitutional isomers with the formula C₆H₁₂O₆. Stereoisomers, on the other hand, have the same connectivity but differ in the spatial arrangement of their atoms.
- D and L Designation: This fundamental classification in stereochemistry is determined by the configuration of the chiral carbon farthest from the anomeric (carbonyl) carbon. In a Fischer projection, if the hydroxyl (-OH) group on this carbon point ot the right, it is a D-isomer. If it points to the left, it is an L-isomer. Most biologically important carbohydrates are D-isomers.

- Enantiomers: These are stereoisomers that are non-superimposable mirror images of each other. The D and L forms of a sugar are enantiomers. For example, D-glucose and L-glucose are enantiomers.
- Diastereomers: These are stereoisomers that are not mirror images of each other. This occurs in molecules with multiple chiral centers when they differ in configuration at some, but not all, of those centers.
- Epimers: All epimers are also diastereomers, but not all diastereomers are epimers. Epimers are a specific type of diastereomer that differ in configuration at only one chiral center. Understanding the epimeric relationships between common hexoses is crucial for the MCAT
- D-mannose is the C2 epimer of D-glucose
- D-galactose is the C4 epimer of D-glucose.
- It is important to note that D-mannose and D-galactose are not epimers of each other; they differ at two chiral centers (C2 and C4) and are therefore classified as diastereomers.
1.3 Cyclic Structures and Anomers
In aqueous solutions, monosaccharides with five or more cabons readily cyclize. This intramolecular reaction occurs when a hydroxyl group attacks the carbonyl carbon. An aldose cyclizes to form a hemiacetal, while a ketose forms a hemiketal.
This cyclization results in two primary ring structures:
- Pyranose: A six-membered ring
- Furanose: A five-membered ring.
Upon cyclization, the former carbonyl carbon becomes a new chiral center called the anomeric carbon. This gives rise to two new stereoisomers, called anomers, which are designated α (alpha) and β (beta). Anomers are epimers that differ in configuration specifically at this anomeric carbon.
The distinction, shown in Haworth projection of a D-sugar, is as follows:
- In the α-anomer, the hydroxyl group on the anomeric carbon is trans to the CH₂OH group (it points down).
- In the β-anomer, the hydroxyl group on the anomeric carbon is cis to the CH₂OH group (it points up).
In solution, the α and β anomers can spontaneously interconvert in a process called mutarotation. This occurs because the hemiacetal ring can open back to its linear form and then re-close, allowing the configuration at the anomeric carbon to change.
1.4 Important Monosaccharide Derivatives
Monosaccharides can be chemically modified to form derivatives with specialized biological roles. Key derivatives for the MCAT include:
- Phosphosugars: A hydroxyl group is replaced by a phosphate group through phosphorylation. Phosphorylated sugars like glucose 6-phosphate and fructose 1,6-bisphophate are critical intermediates in metabolic pathways like glycolysis, trapping the sugar inside the cell.
- Deoxy Sugars: A hydroxyl group is replaced by a hydrogen atom. The most important example is 2-deoxyribose, which forms the sugar-phosphate backbone of DNA.
- Oxidized Sugars: Oxidation of an aldehyde group yields an aldonic acid (e.g., 3-phosphoglycerate), while oxidation of the final carbon yields a uronic acid (e.g., glucuronic acid). These are named with the suffix “-onic acid” or “-uronic acid.”
- Reduced Sugars (Sugar Alcohols): The aldehyde group of an aldose is reduced to a primary alcohol. Examples include glycerol, a component of triacylglycerides, and ribitol.
- Amino Sugars: A hydroxyl group is replaced by an amino group (-NH₂), which is typically protonated and positively charged in the cell. Examples include glucosamine and its derivative N-acetylglucosamine (GlcNAc), which are components of complex polysaccharides.
These fundamental monosaccharide units are linked together to form the more complex carbohydrates that are prevalent in biological systems.
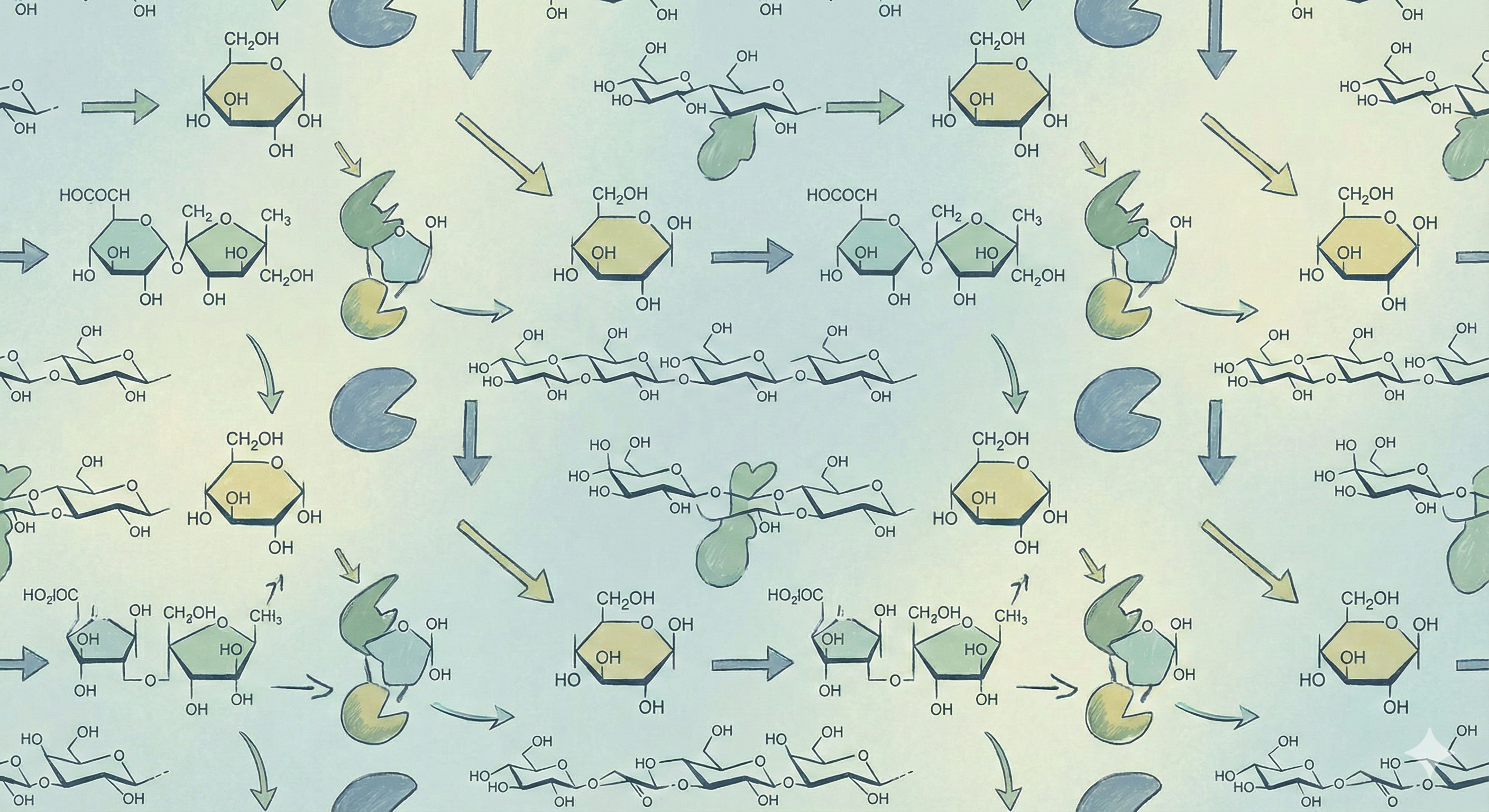
2. Linking Sugars – Complex Carbohydrates
While monosaccharides are the basic units of energy, most carbohydrates in biological systems exist as polymers. The strategic importance of this section is to understand how the glycosidic bond allows for the formation of disaccharides for transport and massive polysaccharides for energy storage and structural integrity.
2.1 The Glycosidic Bond
A glycosidic bond is a covalent bond formed in a condensation (dehydration) reaction between the anomeric carbon of one sugar and a hydroxyl group on another molecule. When this bond forms, the hemiacetal or hemiketal of the first sugar is converted into a more stable acetal or ketal, respectively. This conversion is crucial because it “locks” the anomeric configuration (α or β) of that sugar unit, preventing the ring from opening and thus halting mutarotation.

The standard nomenclature for a glycosidic bond specifies both the anomeric configuration and the carbons involved. For example, a linkage between the anomeric carbon (C1) of a glucose in its α configuration and the C4 hydroxyl group of another glucose is called an α-1,4-glycosidic bond.
Reducing vs. Nonreducing Sugars
This concept relates to the presence of a free anomeric carbon.
- A reducing sugar is any sugar that has a free hemiacetal or hemiketal end. This free anomeric carbon allows the ring to open, exposing an aldehyde group that can be oxidized in laboratory tests like the Tollens test.
- A nonreducing sugar has its anomeric carbons locked in glycosidic bonds, preventing the ring from opening. Sucrose and trehalose are common examples.
- Polysaccharides have polarity: they possess a single reducing end (the one unit with a free anomeric carbon) and one or more nonreducing ends.
2.2 Key Disaccharides for the MCAT
| Disaccharide | Composition & Linkage | MCAT-Relevant Notes |
|---|---|---|
| Maltose | Glucose + Glucose (α-1,4-glycosidic bond) | An intermediate in starch digestion |
| Lactose | Galactose + Glucose (β-1,4-glycosidic bond) | Known as milk sugar. Its β-1,4 linkage requires the specific enzyme lactase for digestion. A deficiency in this enzyme leads to lactose intolerance. |
| Sucrose | Glucose + Fructose (α-1,β-2-glycosidic bond) | Common table sugar. It is a nonreducing sugar because the anomeric carbons of both monosaccharides are involved in the glycosidic bond. |
2.3 Key Polysaccharides for the MCAT
Polysaccharides are large polymers of monosaccharides used for energy storage and structure.
Starch (Plant Energy Storage)
Starch is a polymer of α-D-glucose and is the primary form of energy storage in plants. It exists in two forms:
- Amylose: A linear, unbranched polymer composed of glucose units joined by α-1,4-glycosidic linkages.
- Amylopectin: A branch polymer. It has a main chain of α-1,4 linkages with branch points created by a α-1,6-glycosidic linkages occurring approximately every 25 glucose units.
Glycogen (Animal Energy Storage)
Glycogen is the main form of glucose storage in animals, found primarily in the liver and skeletal muscle.
- Its structure is very similar to amylopectin, with α-1,4 main chains and α-1,6 branch points. However, glycogen is more highly branched, with α-1,6 branch points occuring approximately every 10-14 glucose units, compared to every ~25 units in amylopectin.
- This extensive branching is physiologically significant: it creates numerous nonreducing ends, allowing enzymes to rapidly release many glucose units simultaneously when energy is needed.
Cellulose (Plant Structural Component)
Cellulose is a major structural component of plant cell walls, giving wood and cotton their strength.
- Like starch and glycogen, it is a linear polymer of glucose. However, the critical difference is that the glucose units are joined by β-1,4-glycosidic bonds.
- This stereochemical difference has profound consequences: mammalian digestive enzymes cannot hydrolyze β-1,4 linkages, making cellulose indigestible fiber for humans. The β-linkages allow the polymer to form long, straight, fibrous chains that are ideal for a structural role.
With an understanding of carbohydrate structure, we can now turn to how these molecules are broken down to power the cell.

3. Carbohydrate catabolism – Extracting Energy from Glucose
This section tells the central story of energy extraction in the cell. The oxidation of glucose is a multi-stage process meticulously designed to capture chemical energy in the form of ATP and the reduced electron carriers NADH and FADH₂. For the MCAT, understanding the cellular location, key regulatory points, and net products of each stage is critical.
3.1 Glycolysis: The Universal First Step
Glycolysis is a sequence of ten reactions that oxidizes one molecule of glucose into two molecules of pyruvate. Its key characteristics are:
- Location: Occurs in the cytoplasm of all cells.
- Oxygen Requirement: Does not require oxygen (anaerobic).
- Net Reaction: Glucose + 2 ADP + 2 Pi + 2 NAD⁺ → 2 Pyruvate + 2 ATP + 2 NADH + 2 H₂O + 2 H⁺
Two key regulatory enzymes control the rate of glycolysis:
- Hecokinase: Catalyzes the first step, the phosphorylation of glucose to glucose-6-phosphate. This step traps glucose in the cell and is subject to feedback inhibition by its product, glucose-6-phosphate.
- Phosphofructokinase-1 (PFK-1): Catalyzes the phosphorylation of fructose-6-phosphate to fructose-1,6-biphophate. This is committed step of glycolysis; its product can only proceed through the glycolytic pathway. This reaction is practically irreversible and is the main control point. PFK-1 is allosterically inhibited by high levels of ATP, signaling that the cell has sufficient energy.
3.2 Fermentation: The Anaerobic Fate of Pyruvate
Under anaerobic conditions (i.e., in the absence of oxygen), the electron transport chain cannot function to reoxidize NADH. The purpose of fermentation is to regenerate NAD⁺ from the NADH produced during glycolysis, thereby allowing glycolysis to continue producing a small amount of ATP.
- Lactic Acid Fermentation: In human muscle cells during strenuous exercise, pyruvate is reduced to lactate to regenerate NAD⁺.
- Ethanol Fermentation: In yeast, pyruvate is converted to ethanol and CO₂ in a two-step process that regenerates NAD⁺
3.3 The Pyruvate Dehydrogenase Complex (PDC)
Under aerobic conditions, pyruvate is transported from the cytoplasm into the mitochondrial matrix. Here, the pyruvate dehydrogenase complex (PDC) converts it into acetyl-CoA. This reaction is an oxidative decarboxylation:
- The 3-carbon pyruvate is oxidized to a 2-carbon acetyl unit.
- One molecule of CO₂ is released.
- One molecule of NADH is produced per pyruvate.
- The complex requires the cofactor thiamine pyrophosphate (TPP), which is derived from vitamin B₁.
3.4 The Kerbs Cycle (Citric Acid Cycle or TCA Cylce)
The Krebs Cycle occurs in the mitochondrial matrix and is the central hub of cellular metabolism. Its primary function is to take the 2-carbon acetyl unit from acetyl_CoA and completely oxidize it to CO₂, capturing the released energy in electron carriers.
- The cycle begins when acetyl-CoA combines with the 4-carbon molecule oxaloacetate to form the 6-carbon molecule citrate.
- Through a series of reactions, the cycle regenerates oxaloaetate.
- The net products from one turn of the cycle (from one acetyl-CoA) are: 3 NADH, 1 FADH₂, 1 GTP (or ATPP), and 2 CO₂.
- Since one molecule of glucose produces two molecules of acetyl-CoA, the total yield from one glucose is : 6 NADH, 2FADH, and 2 GTP.
3.5 Electron Transport Chain (ETC) and Oxidative phosphorylation
This is the final stage of cellular respiration and the site where the vast majority of ATP is produced. Its location is the inner mitochondrial membrane. The process has two main goals:
- Reoxidize the electron carriers NADH and FADH₂ back to NAD⁺ and FAD so they can be reused in other pathways.
- Use the energy released from this oxidation to generate a large amount of ATP.

The process unfolds as follows:
- Electron Transport: High-energy electrons from NADH and FADH₂ are passed down a series of protein complexes (cytochromes). As electrons are transferred, energy is used to pump protons (H⁺) from the mitochondrial matrix into the intermembrane space.
- Final Electron Acceptor: Oxygen (O₂) is the final electron acceptor. It accepts the electrons and protons to become reduced to water (H₂O).
- The Proton Gradient: The active pumping of protons creates an electrochemical gradient across the inner mitochondrial membrane, known as the proton-motive force.
- ATP Synthase: This large enzyme complex harnesses the energy stored in the proton gradient. As protons flow back down their concentration gradient into the matrix through a channel in ATP synthase, the energy is used to drive the phosphorylation of ADP to ATP.
3.6 Summary of ATP Yield
The complete aerobic oxidation of one molecule of glucose yields a substantial amount of ATP. The theoretical maximum yield is summarized below.
| pathway | Net Products | ATP Equivalent (Eukaryotes) |
|---|---|---|
| Glycolysis | 2 ATP, 2 NADH (cytosolic) | 2 ATP, ~3 ATP (via shuttle) |
| PDC (x2) | 2 NADH (matrix) | 5 ATP |
| Krebs Cycle (x2) | 2 GTP, 6 NADH, 2 FADH₂ | 2 ATP, 15 ATP, 3 ATP |
| Total | 30 ATP |
4. Anabolism and Alternate Pathways
Metabolic flexibility is key to survival. The body must be able to synthesize glucose when it’s not available from the diet (gluconeogenesis), store it for later use (glycogenesis), and divert its intermediates for the synthesis of other critical biomolecules (pentose phosphate pathway).
4.1 Gluconeogenesis: Synthesizing New Glucose
- Location: Occurs primarily in the liver.
- Function: It is essentially “glycolysis in reverse,” but it is not a simple reversal. To overcome the three thermodynamically irreversible steps of glycolysis, gluconeogenesis emplys four unique enzymes:
- Pyruvate Carboxylase and PEPCK work together to bypass Pyruvate Kinase.
- Fructose-1,6-bisphosphatase bypasses Phosphofructokinase-1 (PFK-1).
- Glucose-6-phosphatase bypasses Hexokinase.
- Energy Cost: Gluconeogenesis is an energetically expensive process, requiring six high-energy phosphate bonds (4 ATP and 2 GTP) to convert two molecules of pyruvate back into one molecule of glucose.
4.2 Glycogen Metabolism: Storage and Release
Glycogenesis (Synthesis)
Glycogenesis is the process of synthesizing glycogen for storage.
- The pathway begins with glucose-6-phosphate, which is converted to glucose-1-phosphate.
- Glucose-1-phosphate is then activated by coupling with UTP to form UDP-glucose.
- The key enzyme glycogen synthase adds this activated UDP-glucose unit to the growing glycogen chain, forming an α-1,4 bond.
Glycogenolysis (Breakdown)
Glycogenolysis is the breakdown of stored glycogen to release glucose.
- The key enzyme is glycogen phosphorylase, which removes one glucose unit at a time from a nonreducing end of the glycogen polymer.
- This process, called phosphorolysis, yields glucose-1-phosphate, which can be converted to glucose-6-phosphate for use in metabolism.
Tissue-Specific Role of Glycogen
- Liver: The liver’s primary role is to maintain blood glucose homeostasis. It breaks down its glycogen stores and, because it contains the enzyme glucose-6-phophatase, can release free glucose into the blood stream for other tissues to use during fasting.
- Skeletal Muscle: Muscle stores glycogen for its own energy needs during exercise. It lacks glucose-6-phosphatase, so the glucose-6-phosphate produced from glycogenolysis is trapped within the muscle cell and enters glycolysis directly.
4.3 The Pentose Phosphate Pathway (PPP)
The Pentose Phosphate Pathway (PPP), also known as the hexose monophosphate shunt, is an alternative pathway that diverts glucose-6-phosphate from glycolysis to serve two major functions:
- Produce NADPH: This reduced electron carrier is distinct from NADH. NADPH is a critical reducing agent used in anabolic processes, such as fatty acid synthesis, and plays a vital role in protecting cells from damage by reactive oxygen species.
- Produce Ribose-5-phosphate: This five-carbon sugar is the essential precursor for the synthesis of nucleotides (for DNA and RNA) and other coenzymes.
The key regulatory enzyme is glucose-6-phosphate dehydrogenase (G6PDH). This enzyme is allosterically inhibited by its product, NADPH, via negative feedback.
Since these pathways have opposing functions-some break down glucose while others build it up or store it-their activity must be tightly and coordinately regulated.
5. Integrated Regulation of Carbohydrate Metabolism
The critical importance of metabolic regulation cannot be overstated. For the MCAT, understanding how the body balances energy production, storage, and synthesis in response to hormonal signals and cellular energy status is a high-yield topic that intergratees all the concepts previously discused.
5.1 Reciprocal Regulation of Glycolysis and Gluconeogenesis
To prevent the wasteful expenditure of energy, opposing metabolic pathways like glycolysis and gluconeogenesis are subject to reciprocal control. This ensures that one pathway is active while the other is suppressed, preventing a futile cycle where both run simultaneously, resulting only in the net consumption of ATP.
The most important regulatory point is the interconversion between fructose-6-phosphate and fructose-1,6-bisphosphate. This step is controlled by a key allosteric regulator, Fructose-2,6-bisphosphate (F-2,6-BP).
- PFK-1 (the committed step of Glycolysis) is strongly stimulated by F-2,6-BP and AMP (a signal of low energy). It is inhibited by ATP (a signal of high energy).
- Fructose-1,6-bisphosphatase (a key enzyme in Gluconeogenesis) is strongly inhibited by F-2,6-BP and AMP.
5.2 Hormonal Control: Insulin and Glucagon
The pancreatic hormones insulin and glucagon are the primary regulators of blood glucose levels. They exert their control largely by modulating the intracellular concentration of the key regulator, F-2,6-BP.
In a High Blood Glucose State (Fed State)
- Insulin is released from the pancreas.
- Insulin signaling in the liver leads to an increase in the concentration of F-2,6-BP.
- This high level of F-2,6-BP potently activates PFK-1, stimulating glycolysis to process the excess glucose.
- Simultaneously, F-2,6-BP inhibits Fructose-1,6-bisphosphatase, shutting down gluconeogenesis.
- Insulin also stimulates glycogenensis, promoting the storage of excess glucose as glycogen.
In a Low blood Glucose State (Fasting State)
- Glucagon is released from the pancreas.
- Glucagon signaling in the liver leads to a decrease in the concentration of F-2,6-BP.
- The drop in F-2,6-BP removes the stimulation from PFK-1, slowing glycolysis. More importantly, it relieves the inhibition of Fructose-1,6-biphosphatase, thereby stimulating gluconeogenesis to produce new glucose.
- Glucagon also stimulates glycogenolysis, triggering the breakdown of liver glycogen to release glucose into the blood.
Hormonal Regulation of Carbohydrate Metabolism
| state | Dominant hormone | Key Regulator (F-2,6-BP) | Active Pathways |
|---|---|---|---|
| Fed state | Insulin | High | Glycolysis, Glycogenesis |
| Fasting state | Glucagon | Low | Gluconeogenesis, Glycogenolysis |
Conclusion: A Unified View of Carbohydrate Metabolism
Carbohydrates are unequivocally central to cellular energy and structure. Their metabolic pathways-from the simple oxidation of a single glucose molecule in glycolysis to the complex, coordinated hormonal regulation of blood sugar-are part of a tightly integrated and exquisitely responsive system. The cell constantly balances the breakdown of glucose for immediate energy with its synthesis and storage for future needs. A strong grasp of these interconnected concepts, from molecular structure to systemic regulation, is essential for a deep understanding of human physiology and for achieving success on the MCAT.
Leave a Reply